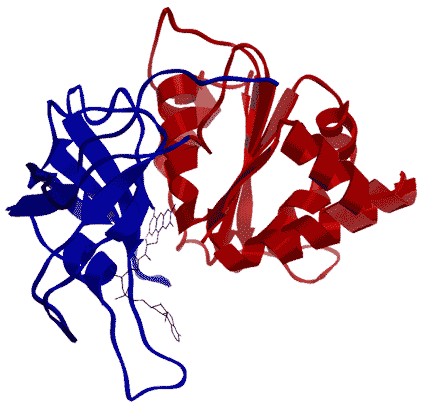
Рисунок Структура молекулы ФНР из Capsicum annuum (в Protein Data Bank структура 1SM4). Рисунок получен с помощью программы Chimera. На рисунке ФАД-связывающий домен обозначен синим цветом, НАДФ+-связывающий – красным, молекула ФАД – серым.
Ферредоксин-НАДФ-редуктаза (ФНР) принадлежит к семейству флавопротеинов (белков, содержащих простетические группы флавинмононуклеотид – ФМН или флавинадениндинуклеотид – ФАД). В качестве простетической группы содержит нековалентно, но прочно связанный ФАД. Содержится у высших растений, эукариотических водорослей и фотосинтезирующих бактерий (Carrillo et al., 1987). ФНР из перца (Capsicum annuum) имеет молекулярную массу 33 кДа. У высших растений ФНР катализирует восстановление НАДФ+ на последней стадии фотосинтетического линейного транспорта электронов и также участвует циклическом транспорте электронов.
ФНР также была обнаружена в разных тканях и организмах, неспособных к фотосинтезу, у которых она участвует в фиксации азота и гидроксилировании стероидов (Arakaki et al., 1997).
При фотосинтезе ФНР катализирует восстановление НАДФ+ до НАДФН в соответствии с реакцией:
Эта реакция может быть разделена на 2 стадии. На первой стадии ФНР катализирует последовательный перенос 2 электронов от 2 молекул восстановленного одноэлектронного переносчика ферредоксина на молекулу ФАД. На второй стадии ФНР использует эти 2 электрона для восстановления НАДФ+ до НАДФН (Carrillo et al., 1987).
|
Рисунок Структура молекулы ФНР из Capsicum annuum (в Protein Data Bank структура 1SM4). Рисунок получен с помощью программы Chimera. На рисунке ФАД-связывающий домен обозначен синим цветом, НАДФ+-связывающий – красным, молекула ФАД – серым. |
Первая структура ФНР была получена с помощью рентгеноструктурного анализа в 1991 году с разрешением 2.6Å (Karplus et al., 1991), а в 1995 разрешение было улучшено до 1.7Å (Bruns et al., 1995; PDB структура 1FNB). Эта была первая структура белка, принадлежащего к семейству флавопротеинов. В 1999 году были получены кристаллические структуры комплекса между НАДФ+ и двумя мутантными ФНР (Deng et al., 1999; PDB структуры 1QFZ и 1QGA).
ФНР состоит из двух доменов, соединенных петлей. Один домен ответственен за связывание простетической группы ФАД – ФАД-связывающий домен (аминокислотные остатки 67–201), другой – НАДФ+-связывающий домен (аминокислотные остатки 202–362). ФАД-связывающий домен образован 6-тяжевым антипараллельным β-бочоноком и одной короткой α-спиралью. НАДФ+-связывающий домен состоит из 5-тяжевого параллельного β-листа, окруженного 6 α-спиралями (Dorowski et al., 2001).
В листьях кукурузы было обнаружено 3 вида ФНР – ЛФНР1, ЛФНР2, ЛФНР3. Эти три изофермента присутствуют в хлоропластах кукурузы примерно в одинаковых концентрациях. Основное различие между изоферментами – это их локализация: ЛФНР1 связана с тилакоидной мембраной, ЛФНР3 – растворимый стромальный фермент, ЛФНР2 присутствует в обеих фракциях. При высоких значениях pH, которые наблюдаются в строме активно фотосинтезирующего хлоропласта, Fd связывается с ЛФНР2 значительно хуже, чем с ЛФНР1, в то время как с ЛФНР3 связывание не на много хуже. Было показано, что количество ЛФНР1, ЛФНР2 увеличивается в условиях повышенного запроса на НАДФН. Наличие нескольких видов ЛФНР позволяет высшим растениям быстро реагировать на изменяющиеся условия (Okutani et al., 2005).
Кроме линейного транспорта электронов, у растений существует циклический транспорт электронов, когда электроны от ФС1 возвращаются к цитохромному b6f комплексу. В результате образуется АТФ без образования НАДФН.
ФНР в хлоропластах высших растений может быть субъединицей цитохромного b6f комплекса (Zhang et al., 2001). Предполагается, что ФНР может быть также связанной с ФС1 (Andersen et al., 1992) и комплексом НАД(Ф)Н дегидрогеназы (Quiles et al., 2000). Bruns и Karplus (Bruns et al., 1995) предположили, что НАДФ+-связывающий домен ФНР может состоять из двух субдоменов. Первый – ответственный за связывание НАДФ+ и второй – содержащий гидрофобный карман, который образует сайт связывания ФНР с мембраной или мембранными белками.
Связанная с цитохромным b6f комплексом или комплексом НАД(Ф)Н дегидрогеназы ФНР может участвовать в циклическом электронном транспорте. Было предложено, что ФНР может регулировать линейный и циклический поток электронов (Bojko et al., 2003). Недавние исследования структуры цитохромного b6f комплекса (Stroebel et al., 2003) выявили новый гемм на стромальной стороне комплекса, который мог бы поддерживать такой циклический транспорт.
Первая структура комплекса Fd и ФНР из листьев кукурузы была получена с помощью рентгеноструктурного анализа в 2000 году с разрешением 2.59Å (Kurisu et al., 2001; PDB структура 1GAQ).
Сайт связывания Fd расположен в углублении между двумя доменами ФНР. Расстояние между редокс центрами в комплексе – 2Fe–2S кластером в Fd и ФАД в ФНР составляет 6.0Å. Взаимодействия между молекулами Fd и ФНР в комплексе в основном электростатические, но вблизи простетических групп – гидрофобные. При образовании комплекса ФНР с Fd конформация ФНР немного изменяется – образуется новая водородная связь, НАДФ+-связывающий домен слегка смещается (Kurisu et al., 2001).